セカンドオピニオンから ペットロスまで どうぶつよろず相談



院長のコラム 2019年6月10日
『ハイエナ③ 系統分類』
 |
 |
|
ハイエナ③ 系統分類 |
||
2019年6月10日 皆様、KVC Tokyo 院長 藤野 健です。 今回はハイエナの系統分類に関する下記論文を採り上げて話を進めます。 Molecular systematics of the Hyaenidae: Relationships of a relictuallineage resolved by a molecular supermatrixKlaus-Peter Koepfli et al.Molecular Phylogenetics and Evolution 38 (3):603-20 , April 2006https://www.researchgate.net/publication/7274943_Molecular_systematics_of_the_Hyaenidae_Relationships_of_a_relictual_lineage_resolved_by_a_molecular_supermatrix(上記 url から全文のPDF が無料で入手可能です)以下、本コラム執筆の参考サイトhttps://ja.wikipedia.org/wiki/ネコ亜目South African large-spotted genetGenetta tigrinahttps://en.wikipedia.org/wiki/Cape_genethttps://ja.wikipedia.org/wiki/マングース科https://en.wikipedia.org/wiki/Banded_mongoosehttps://ja.wikipedia.org/wiki/シママングースhttps://en.wikipedia.org/wiki/Hyenahttps://en.wikipedia.org/wiki/Banded_palm_civethttps://en.wikipedia.org/wiki/Fossa_(animal)https://en.wikipedia.org/wiki/African_palm_civethttps://ja.wikipedia.org/wiki/キノボリジャコウネコhttps://animals.fandom.com/wiki/African_Palm_Civet |
||
 |
 |
|
|
|
ハイエナの系統に関する新しい(とは言っても2006年の)学説を紹介しましょう。下記に引用するのは考察の項の一番最後の部分となります。 Molecular systematics of the Hyaenidae: Relationships of a relictual lineage resolved by amolecular supermatrixKlaus-Peter Koepfli et al. Molecular Phylogenetics and Evolution 38 (3):603-20 , April 20064.4. Inter relationships among the feliform carnivoransOur results show that the clade containing Herpestes, Mungos and Cryptoprocta is placed asthe sister group to the Hyaenidae with 100% nodal support (Fig. 2). Furthermore, we estimate thatthese clades diverged around 29 MYA, in the Middle Oligocene (Table 6). This age is somewhatolder than the 25 MYA age suggested by the hyaenid fossil record (Werdelin and Solounias,1991), but this latter age falls within our estimated credibility interval (Table 6). The joining ofhyaenids and herpestids as sister taxa supports earlier phylogenetic estimates based on analysesof the auditory bulla (Hunt, 1989). Our results are also congruent with previous molecular studiesusing mitochondrial gene sequences (cyt b and ND2) and nuclear gene sequences (intron 1 of thetransthyretin gene and exon 1 of the interphotoreceptor retenoid-binding protein) that sampledonly Crocuta as the solerepresentative of the Hyaenidae (Flynn and Nedbal, 1998; Gaubert andVeron, 2003; Yoder et al., 2003; Yu et al., 2004). The two genera of mongooses represented in ourstudy (Herpestes and Mungos) are classified in the Herpestidae while the Malagasy-endemiccarnivoran Cryptoprocta is traditionally classified as a member of the Viverridae. Our findings,however, confirm the results of a recent study of Malagasy Carnivora that showed thatcarnivorans endemic to Madagascar (traditionally classified in both the Herpestidae andViverridae) are descended from a single common ancestor and share ancestry with theHerpestidae (Yoder et al., 2003). Furthermore, among the four viverrid taxa we sequenced in ourstudy, Nandinia was the most basal lineage within the feliforms (Fig. 2). Collectively, our resultslend additional support to the conclusions of several recent studies that have shown that theViverridae, as traditionally circumscribed, is not monophyletic (Flynn and Nedbal, 1998; Gaubertand Veron, 2003; Yoder et al., 2003; Yu et al., 2004). Overall, the pattern of interrelationshipsamong the feliform families inferred in this study are concordant with these other molecular-based studies, suggesting that diferent regions of the feliform carnivoran genome are tracking thesame phylogenetic history. Although a larger sample of feliform taxa is obviously required tofurther validate these findings, such concordance across diferent studies nonetheless providesconfidence that a stable phylogenetic hypothesis for the primary families of the Feliformia isemerging. |
||
|
|
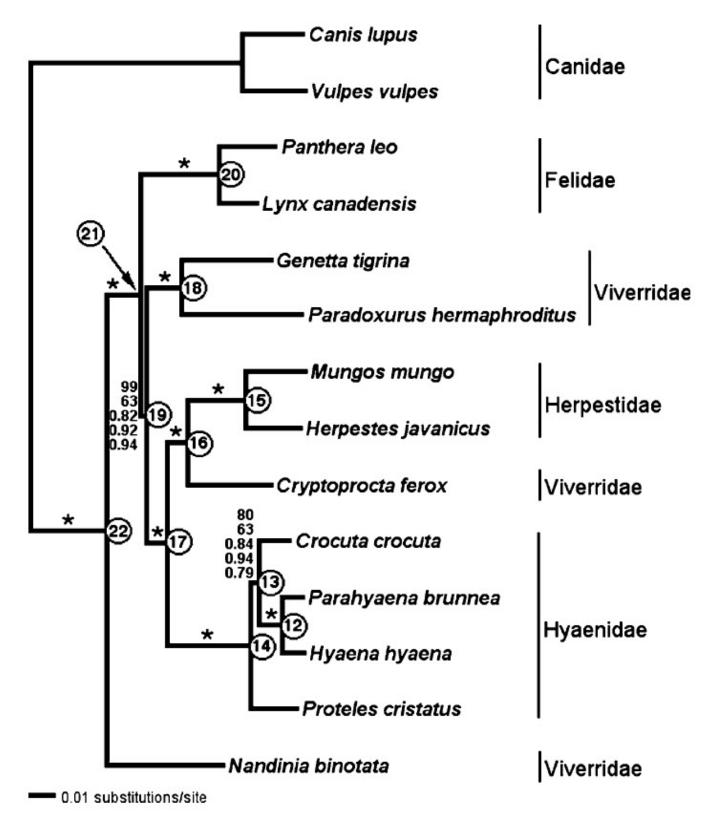
Fig.2 In Molecular systematics of the Hyaenidae: Relationships of a relictual
lineage resolved by a molecular supermatrix Klaus-Peter Koepfli et al.
Molecular Phylogenetics and Evolution 38 (3):603-20 , April 2006
ハイエナ科 Hyaenidae (この図で下から2番目のグループ)は、マングース科
Herpestidae に従来ジャコウネコ科に分類されてきたフォッサCryptoprocta ferox
を加えた新設マングース科 (フォッサをマダガスカルマングース科に独立させる
学説もあります)に姉妹群となるとの結果です(図の17番の節点 nordal で2者は
2900万年前に分岐します)。ジャコウネコ科 Viverridae は実は異なるものの寄
せ集めと判明しましたが (上の図で数カ所に分散しています)、キノボリジャコウネコ
Nandinia binotata(一番下の動物)がネコ亜目の祖先型に近い動物であることが
推測出来ます。
前回コラムで、ハイエナの直接の祖先が、シマヘミガルス(別名タイガーシベット)
Hemigalus derbyanus に良く似た形態を示していたとの説に触れましたが、それが
正しいなら17番まではこの様な胴長のジャコウネコタイプの祖先で推移して来たと
言う話になりますね。ハイエナに枝分かれする際に<一気に>イヌ型化する道を選ん
だと言えます。
上の図で Canidae、Felidae はそれぞれイヌ科、ネコ科です。図で一番上のイヌ科を
除く塊がネコ亜目であり、ネコ科 Felidae、ジャコウネコ科 Viverridae、マングース科
Herpestidae、ハイエナ科 Hyaenidae、から構成されるとされて来ましたが、ジャコウ
ネコ科 Viverridae が実はごちゃ混ぜであることが示唆され、複数に分割して再配
列し直した訳です。これはイヌ科の系統分類の項で触れたジャッカルが、遺伝解析
の結果寄せ集めであったと判明したことに似ています。
|
|
|
(以下院長和訳) 4.4.ネコ型食肉目動物間の相互関係我々の結果は、Herpestes,属 Mungos属及び Cryptoprocta (フォッサ) 属 を含むクレード (分岐群)が節点 nordal から分岐したハイエナ科に対する姉妹群として間違いなく位置づけられる事を示している(図2)。更に、我々はこれらのクレードが2900万年前前後、漸新世中期に分岐したと算定した(表6)。この年代は、ハイエナの化石記録が示唆する年代の2500万年前 (Werdelin and Solounias, 1991) よりは幾らか遡るが、後者の年代は、我々の見積もる信頼区間の内に収まるものである(表6)。ハイエナ科とマングース科を姉妹群として結びつけることは、これらより早期に行われた耳骨胞の分析に基づく研究が提出した、系統発生の推定を支持することになる。我々の結果は、ミトコンドリア遺伝子のシーケンス (cyt b and ND2) 及び遺伝子シーケンス(トランスサイレチン遺伝子のイントロン1 と光受容体間レチノイド結合タンパク質のエクソン1)を用いた以前の分子系統的研究 (Flynn and Nedbal, 1998; Gaubert and Veron, 2003; Yoder et al., 2003; Yu et al., 2004) -これはハイエナ科の代表として僅かに Crocuta属のみをサンプルしたもの- にも矛盾無く一致する。 |
||
|
|

Tamboti Camp, Kruger NP, SOUTH AFRICA 29 January 2015, 19:02
https://upload.wikimedia.org/wikipedia/commons/3/37/Large-spotted_Genet
_%28Genetta_tigrina%29_%2817356502041%29_%28crop%29.jpg
Bernard DUPONT from FRANCE, CC BY-SA 2.0
<https://creativecommons.org/licenses/by-sa/2.0>, via Wikimedia Commons
ケープジェネット CapegenetGenetta tigrina
如何にもジャコウネコ然とした動物です。喜望峰近辺の極く狭い領域に棲息します。

Banded mongoose (M. m. colonus) at Maasai Mara in western Kenya
https://upload.wikimedia.org/wikipedia/commons/0/05/Banded_Mongoose_on_a_log.jpg
Sumeet Moghe, CC BY-SA 3.0
<https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
シママングース Banded mongooseMungos. mungo colonus
如何にもマングース然としたマングースです。

https://www.biolib.cz/IMG/GAL/BIG/345954.jpg
https://www.biolib.cz/en/image/id345954/
キノボリジャコウネコ African palm civet Nandinia binotata
従来はジャコウネコ科に分類されていましたが、現在はキノボリジャコウネコ科として独立
させる見解が強い模様です。紹介した論文の遺伝解析に拠ると、この動物がネコ亜目の
ご先祖に近い系統とされますが、顔は随分とマングース的に感じます。
|
|
|
我々の研究にて科を代表するものとして利用した2つのマングースの属 (Herpestes属 及び Mungos属) は、マングース科に分類されるが、一方、マダガスカル島特有の食肉目動物 であるフォッサは、伝統的にジャコウネコ科の仲間として分類されている。しかしながら我々の研究結果は、マダガスカルの食肉目に関する最近の研究結果、即ち、マダガスカル特有の食肉目 (これらは伝統的に マングース科とジャコウネコ科の両者に分類される) が一つの共通祖先からの子孫であって、祖先をマングース科と共有する (Yoder etal., 2003)、との考えを確かなものとする。更に、我々の研究でシーケンスした4つのジャコウネコ科の分類群の間で、キノボリジャコウネコ属 は、ネコ亜目の中で最も基礎となる系列であった。以上をまとめると、我々の結果は、伝統的にその周囲を境界線で囲まれていたジャコウネコ科が、実は単系統ではないことを示してきた最近の研究 (Flynn and Nedbal, 1998; Gaubert and Veron, 2003; Yoderet al., 2003; Yu et al., 2004) を支持するものとなる。全体的に見れば、本研究で推論されるネコ亜目の科の間の関係のパターンは、これらの他の分子ベースの研究に調和するが、これはネコ亜目の食肉目ゲノムの異なる領域が、同一の系統発生の歴史をそれぞれ別個に辿っていることを示唆する。これらの知見を更に検証する為には、ネコ亜目分類群のより大きなサイズの資料が必要要である事は明らかだが、異なる研究の間でその様な一致が見られることは、それでもなお、ネコ亜目の系統発生上の第一的な科は何であるのか、に関する揺らぎなき仮説が出現しつつあることに、信頼を与えるものである。(以上) |
||
|
|

The fossa has a cat-like appearance, resembling a small cougar
https://upload.wikimedia.org/wikipedia/commons/3/31/Cryptoprocta_ferox.jpg
Chad Teer, CC BY 2.0 <https://creativecommons.org/licenses/by/2.0>, via Wikimedia Commons
フォッサ fossa Cryptoprocta ferox マダガスカル島の沿岸域のみに棲息します。

Madagascar Natural World Date 11 October 2016, 20:56:33
Source https://www.flickr.com/photos/rod_waddington/30256108085/
Author Rod Waddington
https://commons.wikimedia.org/wiki/File:Fossa,_Madagascar_(30256108085).jpg
図の変更無し、ライセンス情報は上記url 内に記述されています。
紹介した論文はマングース科がハイエナ科に一番近いとしています。写真はマダガスカル島に
棲息する大型のマングース、フォッサCryptoprocta ferox。この動物は従来はジャコウネコ科
に分類されてきましたが著者等はマングース科に再分類しています。しかしながら、ハブに噛み
付くようないわゆるマングースとは趣がだいぶ違う様にも感じます。学者によってはこの動物を
マングース科から独立させ、マダガスカルマングース科に置く者もいます。従来ジャコウネコ科に
分類されてきた動物も姿が似たものの寄せ集めだった可能性があるとの話です。このフォッサは
ハイエナにより近い仲間ですが、顔付きがマングースよりは実際ネコに近いと感じます。ただ、
まだネコよりは吻が幾らか伸び出ているようですね。思わずキャメロン監督のアバターが頭に
浮かびました。何とも不思議な雰囲気の動物ですね。
|
|
|
この論文は、遺伝学的解析と形態学的解析を併せて考えると、ハイエナ科はネコ亜目の中でも、ジャコウネコ科ではなく、マングース科に系統的に一番近く、2900万年前に共通祖先から分かれ出た兄弟姉妹の関係にあるとの結論です。また、従来のジャコウネコ科動物の分類には問題があり、血縁関係の距離が異なる様々な動物が紛れ込んでいるとの指摘ですね。これはイヌ科の系統分類の項で触れたジャッカルが、遺伝解析の結果寄せ集めであったと判明したことを想起させます。形態のみに拠る系統分類の弱さが露呈したとも言えるでしょう。 従来のジャコウネコ科動物からはフォッサを分離して、これが寧ろマングースに近いものに置き、更にキノボリジャコウネコの仲間を分離してネコ亜目の最初の祖先型に近いものに置くとの推論です。フォッサに関しては、いわゆるネコに近い位置に置く考えも依然として残っています。まぁ、顔つきから判断すれば確かにネコに近い動物に見え、これがマングースに近いとの遺伝解析結果には、形態学者としては素直に従うには正直躊躇します。 マングースとハイエナとは外見的には体型からしてもあまり似ていない様に感じますが、遺伝解析に基づく系統分類を認めるとすれば、共通祖先から分岐して後に、共通のネコ的要素をベースにしてそれぞれが進化的な改変を受けてきたと考える他はありません。ハイエナの直接の祖先が、シマヘミガルス (別名タイガーシベット) Hemigalus derbyanus に良く似た形態を示していたとの説がありますが、それが正しいなら、胴長のジャコウネコタイプの祖先でマングースとハイエナが分岐する直前まで推移して来たと言う話になります。ハイエナに枝分かれする際に<一気に>イヌ型化する道を選んだと言えますね。 矢張り、ハイエナは相当にイヌ化している様に感じますが如何でしょうか?。他方、ネコ科自体は、ネコ亜目のご先祖様 (キノボリジャコウネコに近い動物とされる)よりも吻の長さを短縮化させている可能性もあり、顔面の平坦化が進行している様にも見えます。こちらは、ネコ亜目の他の仲間からは初期に分岐して大きな発展を遂げています。 ジャコウネコ周囲の動物の系統分類に関しては web 等を検索しても定見を見ているとは言いがたく、今回ご紹介した論文が将来的に受け入れられ続けるのかの保証も有りません。遺伝学に基づく解析は、解析が容易な一部の遺伝子の解析に基づくものであったり、また遺伝子には役立たずの昼行灯の様な遺伝子もあれば、モノを言う遺子もありますので、機械的に比較する方法にどの程度の生物学的な意味があるのか、確かでは無いところも含まれます。系統分類に一つの刺激を与える報告として読むと良いでしょう。 次回最終回の④では、ハイエナの形態的特徴と行動特性を、ジャコウネコやマングースを含めた他のネコ亜目と比較しつつ話を展開します。 |
||
|
|